
Н ОВЫЕ ПУБЛИКАЦИИ РЕСУРСА 07 .07.2026 День Памяти. 7 июля. День памяти моего Отца. 01 .07.2026 Публикации о LCF в 2026 году (Июнь). Статьи и к ниги с упоминанием LCF опубликованные в июне 2026 года. 25 .06.2026 1803LarreyDJ . Автор описывает экзартикуляцию в тазобедренном суставе и методику пересечения LCF , которую именует «межсуставная связка». 16 .06.2026 1948EpsteinI . Реда ктор комментирует слова раввина Самуила (Шмуэля) в трактате Хуллин Вавилонского талмуда, поясняющего расположение LCF по отношению к суставу и ее отличие от седалищного нерва. 1753AstrucJ . Автор, анализируя книгу «Бытие» приводит текст на французском языке с упоминанием травмы тазобедренного сустава, повреждении LCF и последствиях инцидента. 29 .05.2026 Публикации о LCF в 2026 году (Май). Статьи и к ниги с упоминанием LCF опубликованные в мае 2026 года. 28 .05.2026 Интернет-журнал "О КРУГЛОЙ СВЯЗКЕ БЕДРА", май 2026 26 .05.2026 20c.Wikstro...

Статья Strayer Jr LM. The embryology of the human hip joint. (Эмбриология тазобедренного сустава человека, 1943). Автор подробно описывает эмбриональное развитие, прикрепление и строение ligamentum capitis femoris (LCF). Оригинал на английском доступен по ссылке: 1943StrayerJrLM.
ЭМБРИОЛОГИЯ ТАЗОБЕДРЕННОГО СУСТАВА ЧЕЛОВЕКА
ЛЮТЕР М. СТРЕЙЕР, МЛАДШИЙ
Эта статья возникла из потребности в более точных знаниях о развитии нормального тазобедренного сустава человека, чем те, которые можно было бы получить из литературы и учебников по эмбриологии. Было обнаружено, что, несмотря на превосходную работу Bardeen (1,3) в 1901 и 1905 годах, в текстах и статьях (10, 21), опубликованных в 1933 и 1935 годах, можно найти неверные представления о деталях развития. Иллюстрации и фотографии имеющихся моделей не показывают деталей. Поэтому было сочтено, что обзор предмета и представление микрофотографий будут иметь ценность. Наблюдения также были сделаны в надежде соотнести нормальные процессы развития с числом патологических поражений бедра, в которых врожденный фактор считался этиологически значимым.
Подробно описывается формирование вертлужной впадины, головки и шейки бедренной кости, круглой связки, удерживателя Weitbrecht, суставной губы, синовиальной мембраны и поперечной связки вертлужной впадины. Кратко описывается проникновение кровеносных сосудов в головку и шейку бедренной кости и предполагаемая предрасположенность головки бедренной кости к вывиху у одного эмбриона. Иллюстрации эмбрионов размером от 6,75 до 45 мм были выбраны из превосходного материала, имеющегося в Эмбриологической коллекции Майнота Гарвардской медицинской школы. Шесть эмбрионов размером 45 мм, 70 мм, 81 мм, 90 мм, 167 мм и 237 мм соответственно были разрезаны на серийные срезы в Лаборатории патологии Детской больницы в Бостоне. Наблюдения за кровеносными сосудами и последующим развитием были сделаны из этих срезов. В качестве стандарта использовался показатель «темя-копчик», рассчитанный Keibel и Mall.
Литература
Работая над эмбриологией конечностей, Kolliker (11) в 1861 году сделал заявление о том, что «основа всех костей конечностей возникает сначала из одного единого недифференцированного «тела». Когда хрящевое образование впервые начинается во всей этой массе, оно организуется в несколько частей, так что оно разовьется в столько же отдельных хрящей или костей». Это простое и убедительное утверждение является сутью скелетной эмбриологии. Теперь мы знаем, что «единое недифференцированное тело» включает в себя все элементы соединительной ткани конечности.
Многое из написанного о развитии бедра получено из сравнительной анатомии. Bland-Sutton (22) пришел к выводу из таких исследований, что ligamentum teres femoris представляет собой остатки внесуставной связки или мышцы, относящейся к группе приводящих мышц, которая постепенно заключалась внутри сустава по мере того, как бедренная кость становилась все более приведенной и приобретается прямая осанка. Moser (17) проиллюстрировал тазобедренный сустав человеческих эмбрионов размером 30, 34 и 47 мм и, в отличие от утверждения Bland-Sutton, сделал правильный вывод о том, что независимо от процессов развития у животных, у человека эта связка развивается in situ; также суставная щель формируется позади и между связкой и головкой бедренной кости в то же время, что и в других местах сустава. Keith (10) представляет свою концепцию процесса следующим образом: «Круглая связка, лучший пример внутрисуставной связки, появляется у человеческого плода как часть капсулы сустава; у рептилий эта фетальная форма сохраняется. Круглая связка изолируется во время развития головки бедренной кости, которая расширяется как крылья с каждой стороны круглой связки и путем слияния крыльев изолирует ее от капсулы. Отраженная связка на нижней поверхности шейки является частью капсулы, с которой круглая связка была непрерывна». Хотя цитируя Keith, Walmsley (23) находится в прямой оппозиции к Keith, когда он заключает: «У человеческого эмбриона круглая связка полностью свободна при первом появлении суставной щели... Мы считаем, что эти факты указывают на то, что нижний бедренный retinaculum (Weitbrecht) не представляет собой, по крайней мере в целом, сохраняющиеся остатки круглой связки, но что эта брыжейка могла бы и, возможно, действительно существует как retinaculum вертлужной жировой прослойки (или гаверсовой железы), которая возникает в связи с экстра-вертлужной частью круглой связки и инкапсулирует кровеносные сосуды, проходящие через вертлужную щель к вертлужной синовиальной прокладке». Согласно нашим наблюдениям, ретинакула Weitbrecht и гаверсова железа не имеют общего происхождения. Parsons (18) поддерживает точку зрения Bland-Sutton.
В современной литературе существует еще одно заблуждение. Hagopoff (6) исследовал два эмбриона овец длиной 20,4 и 20,7 мм и один человеческий эмбрион длиной 25 мм. Имея этот материал в руках, Hagopoff представил диафиз бедренной кости, продвигающийся путем увеличения, чтобы встретиться с тремя элементами таза; после появления головки, на ней моделируется вертлужная впадина, за исключением тех мест, где вертлужные сосуды своим присутствием препятствуют развитию, например, в вертлужной ямке и поперечной вертлужной связке. Он также утверждает, что апофиз лобковой кости или лобковый зачаток остается независимым от других тазовых зачатков, так что практически не принимает участия в формировании вертлужной впадины. Эти ошибочные концепции имеют современных сторонников. Frazer (5) объяснил образование круглой связки следующим образом: «На ранних стадиях в сочленении участвуют только подвздошная и седалищная кости, синовиальная оболочка переходит от них на капсулу, которая прикреплена вокруг их поверхностей. Лобковая кость покрыта этими волокнами и не имеет суставной области. На следующей стадии покрывающие волокна разрушаются, и лобковая кость приобретает суставную поверхность. Она расширяется, и передняя часть исходной седалищной капсулы оказывается, так сказать, между расширяющейся поверхностью и седалищной костью. Эти волокна сохраняются и остаются прикрепленными к седалищной области, но на их поверхности синовиальная полость расширяется, как показано на последней диаграмме, и также присоединяется к более старой полости ниже, проходя между бедренной костью и нижней частью прикрепленной капсулы. Таким образом, синовиальный туннель формируется более широким внизу, где он включает в себя прикрепление волокон, и сужается на своем бедренном конце, где он прикреплен к ямке». Stewart (21) включает три отдельных заблуждения в свои выводы о развитии бедра. Они цитируются здесь под пронумерованными заголовками в его статье. «1. Бедренная кость и os innominatum дифференцируются от общей скелетной закладки мезобластической инвазией . . . 4. Головка отталкивается к вертлужной впадине на почке шейки. 5. Круглая связка образована боковыми разрастаниями прямоугольной головки, пока ее концы не соединятся, оставляя центральную круглую связку и полусферическую головку бедренной кости...».
Авторами, предшествовавшими превосходной работе Bardeen, опубликованной в 1901 и 1905 годах, являются: Hencke и Reyher, (8), которые не дали отчета о развитии тазобедренного сустава, как иногда утверждается, «но описали конечности 18-20-миллиметровых человеческих эмбрионов, включая сочленение в тазобедренном суставе. Они отметили, что не могли наблюдать развитие os innominatum в трех частях. Schulin (20) предоставил первые иллюстрации развития бедра в виде схематических рисунков коронарных сечений эмбрионов 7 и 25 см, детей в возрасте 1 и 6 лет и взрослого человека соответственно. Moser (17), с его интересом, сосредоточенным на круглой связке, проиллюстрировал бедра эмбрионов человека 30, 34 и 47 мм и описал сустав на этих стадиях. Модели и описания Bardeen в его исследованиях развития человеческого скелета из человеческих эмбрионов длиной от 4 до 50 мм выполнены великолепно. Его описание бедра выглядит следующим образом: «Тазобедренный сустав представлен сначала плотной массой склеро-бластемы. Развитие вертлужной впадины путем врастания и слияния отростков подвздошного, седалищного и лобкового хрящей уже было описано.* Хрящевая полость сустава сначала довольно неглубокая. Но расширение хряща в бластематическую ткань, которая проходит от таза через головку бедренной кости, значительно способствует ее углублению со всех сторон, за исключением области вырезки вертлужной впадины.»
* «Пока человеческий
эмбрион растет от 15-20 мм в длину, происходит быстрое развитие тазовых хрящей.
Около головки бедренной кости каждый дает начало пластинчатому отростку.
Слияние этих отростков производит неглубокую вертлужную впадину. Те, что от
подвздошной и седалищной костей, больше, чем от лобковой кости, и сливаются
друг с другом до того, как лобковая кость сливается с ними. Пропорциональные
площади вертлужной впадины, в которые вносит вклад каждый тазовый хрящ,
по-видимому, по существу, такие же, как и те, которые позже предоставляются
соответствующими тазовыми костями, 2/5 седалищной кости, 2/5 подвздошной кости,
1/5 лобковой кости. При росте вокруг тазобедренного сустава каждый из тазовых
хрящей имеет центробежный рост внутри бластемального таза...».
«Сначала полость сустава полностью заполнена плотной бластемальной тканью, в то время как эмбрион растет от 20-30 мм в длину, образование полости начинается в ткани между хрящевым дном вертлужной впадины и головкой бедренной кости. Первая стадия процесса отмечена уплотнением капсулярной ткани, непосредственно граничащей с суставом, и перихондральной ткани, которая на этой стадии покрывает хрящи на их суставных поверхностях, а также в других местах. В области круглой связки фиброзная полоса также дифференцируется от бластемы сустава. Остальная часть ткани становится более рыхлой по текстуре и в конечном итоге рассасывается».
Вышеизложенное — это
все, что студент эмбриологии может иметь в своем распоряжении для справки.
Существует много статей о развитии бедра в плодный период внутриутробной жизни,
некоторые из которых содержат подробные таблицы измерений сочленяющихся
поверхностей. Одним из лучших из этих авторов является Harrenstein (7), который в значительной степени разделяет теорию LeDamany (12, 13, 14) о врожденном вывихе. Однако ни
один из этих авторов не занимался изучением эмбриона, и нет статей, в которых
можно было бы найти подробности эмбриологии бедра.
Наблюдения
Происхождение зачатка конечности. У эмбрионов длиной от 3 до 4 мм на передней и боковой поверхности стенки тела, на уровне поясничного и 1-го крестцового сегментов, появляется небольшой выступ. Он находится гораздо дальше кпереди, чем можно было бы подумать, и находится в тесной связи с целомомом, вольфовым протоком и сзади с сомитами в этой области. Точное происхождение мезодермы, заполняющей этот выступ, не определено. Bardeen говорит, что он, вероятно, возникает из дорсального несегментированного мезобласта. Lewis утверждает (15), что мы точно знаем, что миотомы не участвуют в формировании мышц конечности, хотя они отвечают за мышцы туловища.
Масса в целом содержит все элементы, необходимые для образования скелета, синовиальной оболочки, связок сустава, мышц и их межмышечных перегородок, и сухожилий, и на этой стадии она лишена кровеносных сосудов и нервов.
После начала бластематической стадии каждый зачаток, такой как os innominatum или бедренная кость, следует распознавать как существующий в трех или более видах тканей или стадиях дифференциации одновременно. Принято различать появление ранней скелетной ткани терминами бластема, предхрящ, хрящ и фетальная кость. Некоторые авторы ограничили очертания различных зачатков частями, состоящими из предхряща или хряща. Взяв бедренную кость размером 17-20 мм в качестве примера, стержень состоит из хрящеподобных клеток, в то время как концы кажутся предхрящевыми, а в области вертела видна масса быстро растущей бластемы. Все эти части кости рассматриваются как часть ее зачатка в той же мере, что и любая другая часть. Не следует игнорировать и бластематические части.
Развитие элементов сустава. Первое появление скелетного зачатка в зачатке конечности в области бедра представляет собой группу плотно упакованных клеток в форме «усеченного конуса с косым основанием, которое приложено к боковой поверхности тела» (Bardeen 1). Область будущего тазобедренного сустава в этой ткани ничем не отличается от других клеток — и остается плотной группой клеток, напоминающих исходную бластему, пока и бедренная кость, и безымянная кость не дифференцируются в легко узнаваемую предхрящевую ткань.
Бедренная кость: Верхний конец бедренной кости неотчетлив, пока, начиная с области середины стержня, не происходит увеличение каждой из бластематических клеток в этой области с увеличением цитоплазмы. Постепенно смежные клетки проксимально и дистально претерпевают аналогичные изменения в теле стержня кости, который очерчен более плотными, более мелкими клетками по периферии. В это время он имеет форму гантели, продольная ось которой направлена перпендикулярно вертлужной впадине под измеренным углом 45 градусов к сагиттальной средней плоскости эмбриона. Начиная с этой стадии существует три центра увеличения, по одному для диафиза и двух конечностей. Центральная часть стержня состоит из наиболее дифференцированной ткани на всех стадиях. В контуре коркового вещества кости в центре имеется небольшая выпуклость. Только проксимально и дистально к средней трети имеются сужения в диаметре стержня. В этих точках клетки гораздо более скучены, чем на концах кости. Головка бедренной кости следует за стержнем и предшествует дистальной части в зрелости клеток, ее составляющих. Таким образом, в контурах бедренной кости имеются три группы клеток на разных стадиях развития. На верхнем конце формируются бластематические выступы для вертелов, и эти клетки затем превращаются в хрящ, как и клетки стержня бедренной кости. Механизм углового положения как приведения, так и сгибания бедренной кости в месте ее соединения с шейкой не может быть объяснен морфологическими наблюдениями (Carey 4). Во время ее развития слои уплощенных хрящевых клеток образуются между зрелым фетальным хрящом в центре и незрелым хрящом на каждом конце кости. Костные соли откладываются в дегенерирующем хряще центра диафиза, и определенный 2-3-клеточный слой кортикальной кости откладывается до того, как капилляры из сильно утолщенной надкостницы прорываются (37 мм) через кору на дистальном конце средней трети, чтобы внедриться в хрящевой костный матрикс, неся с собой фибробласты (позже эндост) и кроветворные клетки костного мозга. Следует отметить, что это первое проникновение кровеносных сосудов происходит на уровне питательной артерии во взрослой бедренной кости.
У эмбриона размером 70 мм кровеносные сосуды входят в хрящ головки и шейки бедренной кости. Капиллярные пучки эндотелия, окруженные клетками, напоминающими незрелые клетки соединительной ткани, возникают из области ретинакул Weitbrecht и входят в многочисленные мелкие лакуны. Клетки хряща, ограничивающие лакуны, изменяются в реакции окрашивания, но они не сжимаются больше, чем другие в этой области, и процесс, по-видимому, представляет собой активное растворение и распад клеток, чтобы сформировать проход для сосуда.
Между питательным сосудом стержня и этими вновь введенными сосудами в голове и шейке нет других, входящих под надкостницу, на этой или 167-миллиметровой стадии. Непосредственно перед и одновременно с вхождением кровеносных сосудов в различные части характер клеток хряща в этой области изменяется от многогранной формы к компактным удлиненным веретенообразным клеткам с длинными отростками, переплетенными под углом друг к другу, и к плотному однородному межклеточному веществу без вакуолей. Этот хрящ, отличающийся по внешнему виду от хряща в центре диафиза, шейки и области эпифизарной линии, более дифференцирован, чем тот, который предшествует формированию энхондральной кости, и время этого изменения предполагает, что его внешний вид тесно связан с измененным питанием.
Эти сосуды были продемонстрированы инъекцией и исследованием серийных срезов как часть сосудистого анастомотического кольца вокруг шейки бедренной кости, окруженного retinaculae Weitbrecht. Это сосудистое кольцо в значительной степени снабжается медиальными и латеральными глубокими огибающими ветвями бедренной артерии и менее непосредственно связано с запирательным, верхними и нижними ягодичными сосудами, которые разветвляются на задней и нижней частях капсулы сустава.
Только у одного из шести более крупных эмбрионов (№ 10, 167 мм) показаны сосуды, входящие в головку из круглой связки бедра, хотя калибр сосудов в каждой связке сам по себе довольно большой.
Вертлужная впадина: В момент своего первого появления в виде линии клеток уменьшенной плотности у эмбриона размером 14–15 мм углубление, расположенное проксимальнее головки бедренной кости в безымянной бластеме, неглубокое и имеет форму блюдца, составляя от 65 до 70 измеренных градусов дуги окружности, проведенной с его включением. Это углубление должно быть углублено и замкнуто, чтобы сформировать полную полуокружность в 180 градусов, прежде чем откроется полость сустава. Одним из факторов, участвующих в этом процессе, по-видимому, является давление со стороны головки бедренной кости, на что указывает тот факт, что при 15 мм она отмечена линией клеток уменьшенной плотности, а при 15-22 мм - аналогичной линией увеличенной плотности. При 23 мм линия уменьшенной плотности появляется проксимальнее головки бедренной кости, и начинается дифференциация круглой связки бедра и других капсульных структур, которые защищают сустав, хотя головка только примерно на две трети окружена хрящом. Шесть эмбрионов длиной 17.5-20 мм были изучены с учетом этого наблюдения. Из них три показали, что около половины головки было покрыто предхрящом, а три показали, что около двух третей головки было покрыто хрящом. Гонады этих эмбрионов не были достаточно дифференцированы, чтобы определить пол. На 23-45 мм хрящ подвздошной кости вырастает над головкой бедренной кости, а губа прикреплена к ее краю. Верхняя часть суставной губы на этой стадии намного больше по отношению к окружающей структуре, чем она будет у зрелого эмбриона. На этой стадии она охватывает больше градусов дуги вертлужной впадины, чем на более поздней стадии развития. Следовательно, кажется, что увеличение протяженности элементов вертлужной впадины отвечает за латеральное смещение губы. Это самая важная часть углубления вертлужной впадины, поскольку нижележащие связи прикрепления круглой связки к поперечной связке и губе не покрывают намного большую часть головки бедренной кости на глубине 45 мм, чем они покрывали, когда они присутствовали в виде недифференцированной бластемы.
Самая ранняя дифференциация бластемы в безымянном зачатке начинается в подвздошной кости чуть выше вертлужной впадины* примерно на стадии 15 мм.
* Обратите внимание, что центры окостенения во всех трех костях значительно удалены от этого места.
Подвздошная кость отстает от стержня и головки бедренной кости в дифференциации на всех стадиях, даже до проникновения кровеносных сосудов в хрящ вертлужной впадины, которое происходит сразу после входа сосудов в головку и шейку бедренной кости. Вскоре после этого, близко к вертлужной впадине, аналогичный центр устанавливается в лобковой кости, позже в седалищной кости. Эти три центра становятся предхрящевыми, а затем хрящевыми, так что в оставшейся бластеме мы сначала видим контур Y, как у ребенка. Образование хряща распространяется от этих центров в трех зачатках. Нижняя часть Y проходит через середину вертлужной ямки, которая образована отверстием в боковых границах лобковой и седалищной костей, когда они соединяются (см. рис. 10). Подвздошная и седалищная кости очень быстро сливаются в хряще плода (13.3 мм), чтобы устранить заднее крыло Y. Затем подвздошная и лобковая кости начинают сливаться в хряще, начиная с медиальной и верхней границы и продвигаясь вниз к вертлужной впадине (23 мм). Наконец, лобковая и седалищная кости сливаются от своего внутреннего края наружу, так что при 45 мм все, что остается от исходных бластемальных соединений, представляет собой небольшой угол, открытый латерально на вершине вертлужной впадины. В этой ямке гаверсова железа развивается как масса рыхлой ткани, пронизанной множеством мелких капилляров. Тот факт, что лобковая часть Y остается в виде бластемы в вертлужной впадине до последнего, может быть связано как с соединением этих костей в симфизе (см. рис. 6, 7 и 8), так и с изменениями в направлении костей, необходимыми для достижения этого соединения, как и с развитием вертлужной впадины. В отличие от других авторов, мы обнаруживаем, что часть вертлужной впадины, образованная бластемой и хрящом лобковой кости, остается той же по контуру и размеру по отношению к двум другим хрящам на протяжении всего развития сустава.
Суставная губа, поперечная вертлужная связка: Во время первого предположения об образовании вертлужной впадины наблюдается уплотнение бластемы на периферии углубления, которое представляет собой суставную губу. На 19.3 мм овальные клетки можно увидеть при большом увеличении, расположенные концентрически с ободом вертлужной впадины. Сравнивая относительную массу, занимаемую этими клетками, с массой безымянной бластемы, можно сделать вывод, что расширения клеток не происходит, но что они достигают своего окончательного положения из-за увеличения края подвздошной, седалищной и лобковой костей, в результате чего губа выдвигается над головкой бедренной кости. Внизу у поперечной связки кольцо не поддерживается хрящом медиально, и во время открытия полости сустава поперечная связка не охватывает головку бедренной кости дистальнее ее наибольшего диаметра, так что смещение головки могло бы произойти в этом месте вниз. Сверху губа покрывает наибольший диаметр и далее, образуя прочное защитное покрытие для головки бедренной кости. Эти анатомические особенности приводят нас к предположению, что место поперечной связки тазобедренного сустава является, безусловно, самым слабым местом в его структуре. Это иллюстрируется микрофотографией эмбриона 167 мм, у которого одно бедро было остро согнуто (рис. 18).
Суставная капсула и синовия: В течение периода развития до 45 мм капсула представляет собой рыхло связанную полосу ткани, которая при 22-23 мм может быть различима как слой клеток, лежащих под мышечными зачатками и над суставной губой, и соединяющихся с надхрящницей бедренной кости ниже. Ядра этих клеток, которые имеют округлую или овальную форму в сечении, становятся все более и более веретенообразными, но клетки очень рыхло уплотняются при 44-45 мм, когда можно увидеть небольшое уплотнение, образующее zona orbicularis. Зона четко определяется у 167-мм эмбриона (рис. 17) на полпути вниз по шейке бедренной кости на ее верхней поверхности.
Во время открытия суставной щели невозможно морфологически различить клетку внутреннего края капсулы, которая в конечном итоге сформирует синовиальную оболочку, и те клетки самой капсулы. Это также верно для 167-мм эмбриона. При 237 мм в области расслабленной капсулы можно увидеть определенные кубовидные клетки, но над головкой бедренной кости, вертлужной впадиной и растянутой капсулой они имеют веретенообразную форму.
После открытия полости сустава клетки капсулы вырабатывают больше межклеточного вещества и фибрилл, создавая впечатление увеличения относительной толщины и прочности.
Ligamentum Teres Femoris: На 23 мм обнаруживается первое предположение об упорядоченном расположении клеток в области круглой связки. Отдельные клетки выглядят как примитивные фибробласты со слегка овальными ядрами, плотно прилегающими к головке и на одной линии с длинной осью бедренной кости. Ниже эти клетки смешиваются с клетками поперечной связки. Сагиттальные сечения (рис. 9) показывают треугольный контур клеток, прикрепленных по длине вертлужной связки и сходящихся в ямке. Головка бедренной кости круглая, что можно определить по сечениям, выполненным в сагиттальной, фронтальной и поперечной плоскостях. На стадии нет никаких указаний на углубление в головке для приема круглой связки.
Последующее развитие включает увеличение и небольшое удлинение по мере того, как бедренная кость становится более приведенной. Ее разделение для образования свободной массы внутри сустава происходит одновременно с открытием остальной части полости путем вакуолизации, дегенерации и расщепления между клетками вдоль ее края. Только после этого можно ясно увидеть, что связка должна прикрепляться к медиальному краю вертлужной ямки позади поперечной вертлужной связки.
Три места происхождения связки не одинаково развиты у всех эмбрионов. Седалищная кость и медиальная сторона поперечной связки являются точками происхождения у всех исследованных эмбрионов. Лобковая ножка и прикрепление, проходящее под основанием гаверсовой железы, изменчивы. В секциях двух эмбрионов, показанных на рис. 13 и 15, есть мощная полоса, проходящая от поперечной связки медиально под гаверсовой железой к нижнему краю вертлужной вырезки, которая не присутствовала у пяти других на этой стадии развития. В плоскостях иллюстрированных сечений лобковая и седалищная связки перпендикулярны сечению, а их относительный размер варьируется у каждого эмбриона.
Кровеносные сосуды присутствуют в ткани гаверсовой железы и круглой связке при 23 мм. Как мы показали, ретинакулярные сосуды входят в голову и шейку при 70 мм. Только один эмбрион (167 мм) из семи, исследованных после открытия полости сустава, показал сосуды, входящие в головку из связки. У самого крупного исследованного плода (237 мм) здесь не было никаких сосудов.
Полость сустава: Открытие полости сустава является одновременно дегенеративным и механическим процессом. Имеются наблюдения компетентных исследователей (Reyher, Schulin, Moser, Bardeen), подтверждающие мнение, что никакое врастание ткани извне не обеспечивает выстилку сустава.
Признаки дегенерации видны на 23 мм; наблюдается увеличение межклеточных пространств в области клеток, лежащих между головкой бедренной кости, круглой связкой и вертлужной впадиной. На 36-42 мм, по-видимому, образуются пространства, заполненные жидкостью, где нет противолежащих поверхностей сустава. Изолированные клетки, взвешенные в этой жидкости, демонстрируют автолитическую дегенерацию за счет потери своих клеточных контуров и выцветания своей ядерной части. Другие ядра пикнотичны. Пространства также содержат длинные фибриллы, которые, вероятно, являются результатом дегенерации клеток.
Признаки механического фактора обнаруживаются при наблюдении за тесно прилегающими хрящевыми поверхностями сустава. В этих областях первыми признаками разрыва являются длинные полосы хорошо сохранившихся клеток, протянутые по диагонали от вертлужной впадины до головки бедренной кости. Отростки клеток так прочно прикреплены друг к другу, что, по-видимому, выдерживают значительное натяжение. Расщепление не происходит на одном уровне между дугообразными линиями клеток. Один конец нити, пересекающей пространство, может возникнуть на внешнем слое вертлужной впадины и быть вставленным или прикрепленным к слою 3-4 клеток в глубине головки бедренной кости. Нельзя отрицать, что этот вид может быть артефактом; однако из шести исследованных эмбрионов размером от 36 до 45 мм суставная щель 36 мм (рис. 11 и 12) находится в стадии развития без расщепления клеточных слоев. Все остальные, 37, 42, 44.3 и 45 мм соответственно, показывают расщепление клеток вдоль краев сустава, а также растворение клеток (рис. 13 и 14). На рис. 15 пространство хорошо открыто выше и ниже шейки бедренной кости. Также имеются полоски клеток, лежащих под суставной губой. Все они больше, чем при 36 мм где открытие, по-видимому, является чистым клеточным автолизом. Если бы это был артефакт, можно было бы ожидать, что он будет отмечен больше всего у молодых, наименее развитых эмбрионов.
Предполагается, что,
вероятно, факторы иннервации и функции мышц оказывают некоторое влияние на
время открытия суставной щели, и рано созревающий нервно-мышечный механизм
может привести к раннему открытию сустава, в то время как медленно
развивающийся нервно-мышечный аппарат может позволить эмбриону достичь большей
длины до того, как это произойдет.
Расширение суставной щели внизу по шейке бедренной кости происходит таким образом, чтобы оставить надхрящницу и отраженную капсулу, которая покрывает ретинакулу Weitbrecht. Нет никаких возможных объяснений на основе чисто морфологических наблюдений того, как происходит выбор этой плоскости.
Обсуждение
Эти наблюдения приводят к одному важному выводу, а именно, что все элементы тазобедренного сустава дифференцируются in situ из одной массы мезодермы. Этот момент уже был выдвинут Hencke и Reyher, Moser, Schulin, Walmsley и Bardeen.
Доказательства сравнительной анатомии не могут быть применены к фактическому развитию человеческого бедра. Современные авторы, использующие этот источник в качестве основы для утверждений, — это Keith и Frazer. Утверждения этих авторов касаются формирования круглой связки и роли лобкового зачатка в формировании вертлужной впадины. Bardeen утверждает, и мы подтверждаем это утверждение, что доля тазовых зачатков, входящих в формирование вертлужной впадины, по сути такая же, как у взрослого человека. Путаница существует, потому что авторы не считают бластематические части зачатков как-либо связанными с хрящевыми частями, и рассматривают бластему как некую активно мигрирующую или врастающую синовиальную специальную ткань. По нашим наблюдениям, а также по наблюдениям других (Reyher, Schulin, Moser, Bardeen), синовия не развивается как врастание, а из клеток in situ вдоль линии расщепления, которая появляется между клетками как интимная часть их соответствующих зачатков. Большая часть лобкового зачатка остается в виде бластемы дольше, чем в любой другой лобковой кости. То, что это происходит, может быть связано с закрытием лобкового симфиза. Но, учитывая эти факты, нельзя считать, что она вносит меньший вклад в формирование вертлужной впадины, чем пропорционально ее более поздней части в составе вертлужной впадины. Аналогично мы не можем найти никаких доказательств того, что круглая связка формируется между крыльями, отходящими от головки бедренной кости, как утверждает Keith.
Распространение хряща в бластематические части различных зачатков не может быть объяснено ни в малейшей степени этими наблюдениями, которые избегают динамических интерпретаций.* Наклон стержня изначально прямой бедренной кости для формирования шейки; механические аспекты открытия суставной полости; и расширение тазовых хрящей в бластему вокруг головки бедренной кости близко приближаются к динамическим аспектам эмбриологии, которые рассматривались Carey (4). Тем не менее, кажется неправильным рассматривать верхний конец бедренной кости как почку или отросток (Stewart, Hagopoff). Мы должны просто повторить, что каждая часть развивается in situ из склеробластемы путем непрерывного роста и простого увеличения без грубых изменений во взаимоотношениях.
* Carey писал о динамических и физиологических аспектах эмбриологии, указывая на некоторые очень интересные корреляции.
Нас интересовали расположение и способ проникновения кровеносных сосудов в зачатки, поскольку места и процесс соответствуют некоторым врожденным аномалиям, обнаруженным в области бедра. Большинство клинических случаев врожденного отсутствия бедренной кости показывают отсутствие проксимальных двух третей диафиза бедренной кости. Первое проникновение кровеносных сосудов в диафиз происходит на стыке средней и дистальной третей бедренной кости и в течение длительного периода развития (36-70 мм) представляет собой единственный сосуд в надхрящнице и надкостнице всей кости. На глубине 70 мм кровеносные сосуды входят в головку и шейку бедренной кости из retinacula Weitbrecht в месте поражений, обнаруженных при состоянии так называемой врожденной coxa vara. Эти наблюдения ценны только в свете того, что другие наблюдатели могут добавить к ним. Роль связки круглой кости бедра в снабжении кровью этой области важна при плоском тазобедренном суставе, переломах и инфекциях бедра. Мы, как и многие другие авторы, отметили, что сосуды в этой области различаются по размеру и количеству.
Проблема врожденного
вывиха привлекала многих авторов, но почти никто не обращался к эмбриологии за
ответами на свои вопросы. Возможность наблюдать положение плода на
микрофотографии была случайной, но она ценна тем, что подчеркивает взаимосвязь
головки бедренной кости и вертлужной впадины после открытия полости сустава.
Выводы
Все элементы
тазобедренного сустава дифференцируются in situ в одной массе бластемы.
Головка бедренной
кости имеет шаровидную форму на всех этапах своего развития.
Относительные
пропорции зачатков лобковых костей, входящих в формирование вертлужной впадины,
одинаковы у ранних эмбрионов, на более поздних стадиях и в постнатальной жизни.
Круглая связка не
проявляет никаких признаков возникновения внесуставно или внутри бороздки в
головке бедренной кости у человеческого эмбриона.
Врожденный вывих
бедра не может возникнуть до открытия суставной полости и связан с анатомией
нижней части тазобедренного сустава и положением плода.
ССЫЛКИ
1 Bardeen, C. R.: Studies of the development of the human skeleton. Am. J. Anat., 1905, 4, 265.
2 Bardeen, C. R.: [In] Manual of Human Embryology (Keibel and Mall). 1910, 1, 483.
3 Bardeen, C. R., and Lewis, W. H.: Development of the limbs, body-wall and back in man. Am. J. Anat., 1901, 1, 1.
4 Carey, E. B.: Direct observations on the transformation of the mesenchyme in the thigh of the pig embryo. J. Morphol., 1922, 37, 1.
5 Frazer, J. E. S.: Anatomy of the Skeleton. 3rd ed., London, Churchill, 1933, p. 128.
6 Hagopoff: De l'origine et du mode de developpement embryonnaire de l'articulation de la hanche. Compt. rend. Soc. de biol., 1898, 50, 51.
7 Harrenstein, R. J.: Een kritieke periode in de ontwikkelung van het heupgewricht. Nederl. tijdschr. v. geneesk., 1924, 68, 2328.
8 Hencke, W., and Reyher, C.: Studien ueber die Entwickelung der Extremititen des Menschen. Sitzungsber. d. k. Akad. d. Wissensch., Wien, 1874, 70, 217.
9 Keibel, F., and Mall, F. P. (editors): Manual of Human Embryology. Phila., Lippincott, 1910-1912, 2 v.
10 Keith, Sir Arthur: Human Embryology and Morphology. 5th ed., N. Y., Wood, 1933, p. 515.
11 Kolliker, A. von: Entwickelungsgeschichte des Menschen und der h3heren Thiere. Leipzig, Engelmann, 1861, p. 130. (Quoted by Schulin.)
12 Le Damany, P.: La cavite cotylide. J. de l'anat. et physiol., 1904, 40, 387.
13 Le Damany, P.: Die angeborene Hiuftgelenksverrenkung. Ztschr. f. orthop. Chir., 1908, 21, 129.
14 Le Damany, P.: La luxation congenitale de la hanche. Paris, Alcan, 1912.
15 Lewis, W. H.: The development of the arm in man. Am. J. Anat., 1901, 1, 145.
16 Lewis, W. H.: [In] Manual of Human Anatomy (Keibel and Mall). 1910, 1, 483.
17 Moser, E.: Ueber die Ligamentum teres des Huftgelenks. Morphol. Arb., 1893, 2, 36.
18 Parsons, F. G.: The joints of mammals compared with those of man. Pt. II. The hip joint. J. Anat. & Physiol., 1900, 34, 301.
19 Reyher, C.: On the cartilages and synovial membranes of the joints. J. Anat. & Physiol., 1873-4, 8, 261.
20 Schulin, K.: Ueber die Entwickelung und weitere Ausbildung der Gelenke des menschlichen Korpers. Arch. f. Anat. u. Physiol., Anat. Abt., 1879, 240.
21 Stewart, S. F.: The physiological treatment of congenital dislocation of the hip. J. Bone & Joint Surg., 1935, 33, 11.
22 Sutton, J. B.: The ligamentum teres. J. Anat. & Physiol., 1882-3, 17, 191.
23 Walmsley, T.: A note on the retinacula of Weitbrecht. J. Anat., 1917, 51, 61.
Выражаем искреннюю признательность за полезную критику и поддержку при подготовке данной статьи доктору Фрэнку Р. Оберу, доктору Уильяму Т. Грину, доктору Сидни Фарберу из Детской больницы и доктору Дж. Л. Бремеру из Гарвардской медицинской школы.

Рис. 1. Микрофотография X55 H.E.C. Эмбрион 2300, длина 6,75 мм, слайд 420. Зачаток конечности сформирован. Показаны его связь с сомитами, целомом, вольфовыми дуэтами. Клетки неотличимы друг от друга, за исключением множества митозов. С этого периода зачаток конечности становится отдельным образованием, и смещение каудально и латерально происходит одновременно с удлинением эмбриона в целом.
Рис. 2. Микрофотография X39 H.E.C. Эмбрион 1000, длина 10 мм, слайд 657. Скелетные бластемы сохраняют свой однородный характер. Периферические нервы выросли в пласт из спинного мозга и в безымянной бластеме распадаются на три основные массы, которые проникают до области середины бедра. Мышечные и костные зачатки отдельно не различимы.
Рис. 3. Микрофотография X21 H.E.C. Эмбрион 2051, длина 15 мм, слайд 1010. При высоком увеличении гантелевидная бедренная кость показывает клетки в линиях, перпендикулярных длинной оси, становясь выпуклой на каждом конце. Шаровидная головка бедренной кости не соответствует вертлужной впадине. Отведение бедра на 50° помещает бедренную кость почти под прямым углом, 80° с гнездом, способствуя стабильности соединения.
Рис. 4. Микрофотография X30 H.E.C. Embryo 2155, длина 17,5 мм, слайд 1169. Масса глубоко окрашенных клеток отмечает сустав. Группы мышц очерчены, некоторые сходятся в проекции бластемы, определяя большой вертел. Бедренная кость, покрытая надхрящницей, показывает хрящевые клетки, сжимающиеся от матрикса в центре, и дугообразные столбы предхрящевых клеток, составляющих концы. Элементы безымянных зачатков можно идентифицировать.

Рис. 5. Микрофотография X30 H.E.C. Эмбрион 1597, длина 19.3 мм, слайд 807. Центр бедренной кости показывает зрелый плодный хрящ. Наклон верхнего конца очерчивает шейку, которая образует угол 60° к средней линии и 160° к стержню. Обратите внимание на распределение клеток в поперечных линиях между шейкой и стержнем. Область сустава густо окрашена.
Рис. 6. Микрофотография X21 H.E.C. Эмбрион 2046, длина 23 мм, слайд 1606. Линия сустава представляет собой зону пониженной плотности. Бедренная кость становится приведенной под углом 30-40° к средней линии. Шейка выглядит длиннее и более угловатой на диафизе, 65-70° к средней линии и 150-155° к диафизу. Ложный таз с передней верхней остью подвздошной кости показан как проекция предхряща.
Рис. 7. Микрофотография X30 H.E.C. Эмбрион 913, длина 39 мм, слайд 1064. Найдено отведение бедренной кости на 20-28° к средней линии. Это, по-видимому, происходит за счет сомитного вращения головки в вертлужной впадине и увеличения угла наклона шейки к диафизу бедренной кости. Обратите внимание на хорошо развитую суставную губу.
Рис. 8. Микрофотография X30 H.E.C. Эмбрион 913, длина 30 мм, слайд 1094. Тот же эмбрион, что на рис. 7. Видна бластема в переднем крае Y между лобковой и седалищной. Обратите внимание на расположение центра окостенения в подвздошной кости в отличие от ядра цилиндрификаций около вертлужной впадины на рис. 3 и 6. Ход круглой связки от поперечной связки до ямки выражен. Видны суставная губа и взаимосвязь с капсулой. Лобковый симфиз сближается.

Рис. 9. Микрофотография X55 H.E.C. Эмбрион 1598, длина 28.8 мм, слайд 218. Сагиттальный разрез через медиальную стенку вертлужной впадины показывает относительную пропорцию седалищных и лобковых прикреплений круглой связки и область вертлужной ямки. На этой стадии Y-образное соединение отсутствует, так как сращение в хряще уже произошло. Этот хрящ будет очерчивать Y у ребенка, когда центры окостенения тазовых костей приблизятся к вертлужной впадине. Таким образом, мы видим, что Y дублируется как в бластеме, так и в хряще на разных стадиях, хотя точки начала хондрификации и окостенения не совпадают в тазовых костях, как в бедренной кости.
Рис. 10. Микрофотография X50 II.E.C. Эмбрион 737, длина 22.8 мм, слайд 406. Поперечное сечение показывает углубление между лобковой и седалищной костями, образующее вертлужную ямку. В нем видны начала круглой связки и гаверсовой железы. Антеверсия шейки бедренной кости развилась, образуя угол 30° к средней линии эмбриона. Круглая связка развивается на месте без формирования борозды в головке бедренной кости. Короткие вращательные мышцы бедра хорошо очерчены.
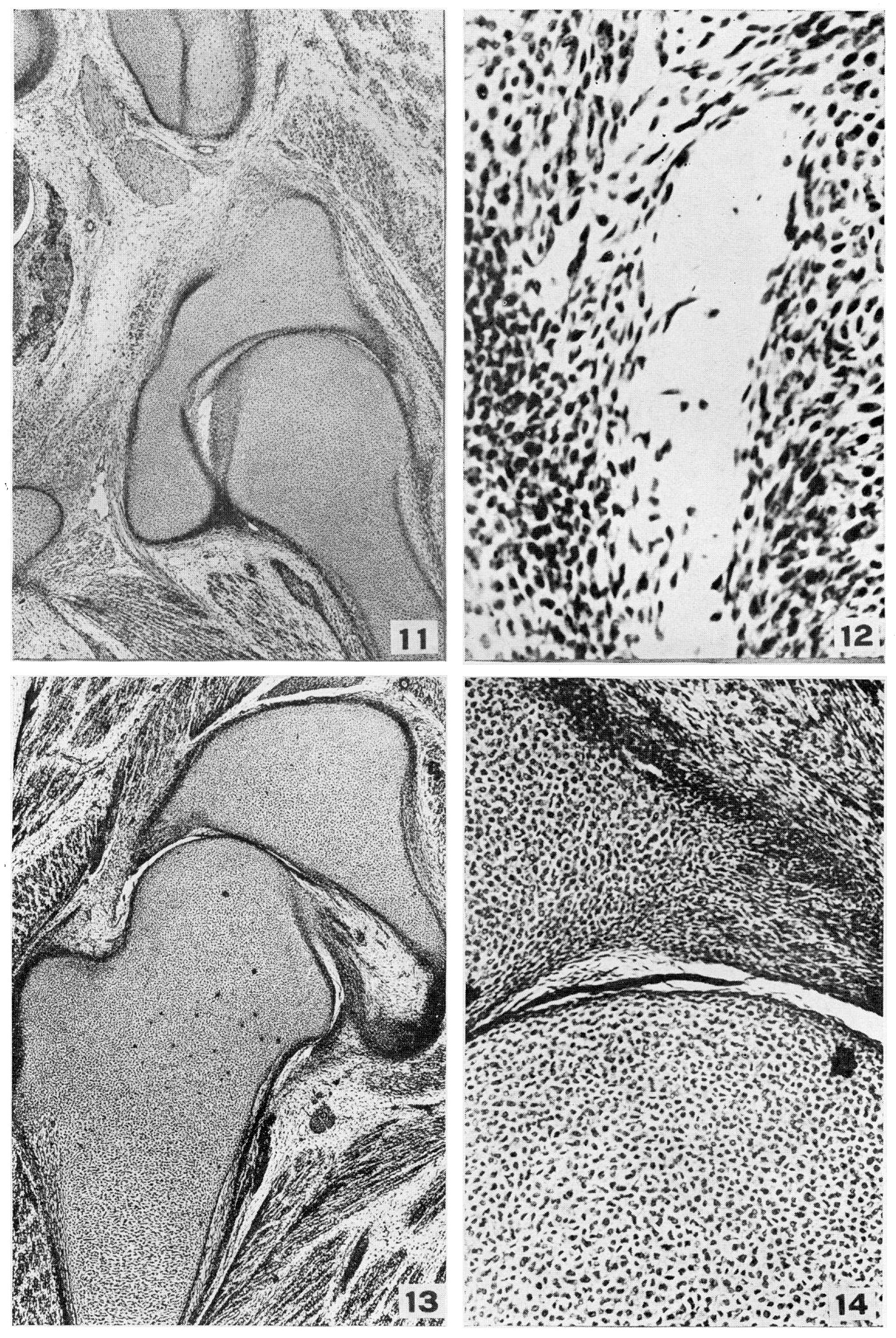
Рис. 11. Микрофотография X20 H.E.C. Эмбрион 2059, длина 36 мм, слайд 2531. Показаны пространства, появляющиеся в ткани вокруг головки бедренной кости. Определены все элементы сустава. Вертлужная впадина, суставная губа и поперечная вертлужная связка образуют 180° окружности. Кровеносные сосуды присутствуют в надхрящнице, суставной капсуле, круглой связке и гаверсовой железе, но ни один из них не присутствует в хряще или стрежне костей.
Рис. 12. Микрофотография X 140 H.E.C. Эмбрион 2050, длина 36 мм, слайд 2531. Значительное увеличение пространства на рис. 11. В пространствах плавают клетки, имеющие нормально выглядящие ядра и длинные фибриллы. Имеются более мелкие клетки с пикнотическими ядрами, а также фрагментированные нематериальные, предполагающие тени ядер. Пространства появляются под круглой связкой, между ней и головкой бедренной кости и внутри капсулы дистальнее суставной губы.
Рис. 13. Микрофотография X20 H.E.C. Эмбрион 838, длина 42 мм, слайд 1140. У двух (рис. 13 и 15) из 6 эмбрионов, исследованных на этой стадии, полоса от круглой связки проходит медиально к нижнему краю ямки вертлужной впадины, а также к поперечной связке, в дополнение к обычной седалищной полосе. Обычно в этой точке имеется только тонкая полоса капсулы, отходящая от лобковой и седалищной кости.
Рис. 14. Микрофотография X140 H.E.C. Эмбрион 838, длина 42 мм, слайд 1140. Высокое увеличение Рис. 13. Пластинки клеток, проходящие через сустав, не показывают признаков дегенерации, как это видно на Рис. 12. Они веретенообразные и того же характера, что и у круглой связки. Синовиальные клетки кубовидной формы отсутствуют.

Рис. 15. Микрофотография X20 H.E.C. Эмбрион 1611, длина 44,3 мм, слайд 2623. Следует отметить полость сустава; многочисленные сосуды в гаверсовой железе, круглой связке и нижней удерживающей связке Weitbrecht; многогранные хрящевые клетки шейки, противостоящие поперечному расположению хряща дистально. Присутствует полость от лобковой кости до круглой связки. Суставная губа и поперечная вертлужная связка выходят за пределы наибольшего диаметра головки бедренной кости.
Рис. 16. Микрофотография на малом увеличении C.H. Эмбрион 2, длина 70 мм, слайд 117. Кровеносные сосуды входят через небольшие лакуны, выдолбленные в головке и шейке. У трех исследованных эмбрионов размером от 70 до 90 мм, сосуды, входящие в нижнюю ретинакулу, кажутся более развитыми, чем те, что из ретинакулярных поверхностей и вертельной ямки. Центр шейки состоит из округлых многогранных клеток. На периферии клетки удлинены и уплощены. Это изменение происходит одновременно с вхождением капилляров. Крупные сосуды показаны в круглой связке.

Рис. 17. Микрофотография на малом увеличении C.H. Эмбрион 10А, длина 167 мм, слайд 155. Фронтальный разрез, показывающий количество сосудов, присутствующих в головке и шейке бедренной кости, которая находится в нормальном внематочном отношении к вертлужной впадине. Показан один сосуд, входящий через круглую связку. Весь хрящ теперь состоит из веретенообразных клеток с плотным гомогенным межклеточным веществом. Синовиальные клетки веретенообразные, уплощенные с несколькими переходными клетками над расслабленными капсульными областями.
Рис. 18.
Микрофотография на малом увеличении C.H. Эмбрион 10. длина 167 мм, слайд 245. Разрез
с бедренной костью в положении сгибания на 90° и внешней ротации. Головка
бедренной кости откинута вниз и латерально, так что ее наибольший диаметр
находится вне окружности суставной губы и вертлужной связки. Нижняя капсула
растянута и выпячена. Если бы сила была приложена по длинной оси бедренной
кости, головка бы вышла за пределы суставной губы, что привело бы к нижнему
вывиху.
Внешние ссылки
Strayer JrLM. The
embryology of the human hip joint. The Yale journal of biology and medicine.
1943;16(1)13-26. pmc.ncbi.nlm.nih.gov
Авторы и принадлежность
LUTHER M. STRAYER, JR. (ЛЮТЕР М. СТРЕЙЕР-МЛАДШИЙ). Отделение
ортопедической хирургии Гарвардской медицинской школы и отделение патологии
детской больницы, Бостон, Массачусетс.
Ключевые слова
ligamentum capitis femoris, ligamentum teres, связка головки бедра, анатомия, эмбриология, развитие, прикрепление, строение
NB! Добросовестная практика использования: копирование для целей критики, обзора, комментариев, исследований и частного изучения в соответствии с Законами об авторском праве: Copyright Laws of the US: 17 U.S.C. §107; Copyright Law of the EU: Dir. 2001/29/EC, art.5/3a,d; Copyright Law of the RU: ГК РФ ст.1274/1.1-2,7.