
Н ОВЫЕ ПУБЛИКАЦИИ РЕСУРСА 01 .07.2026 Публикации о LCF в 2026 году (Июнь). Статьи и к ниги с упоминанием LCF опубликованные в июне 2026 года. 25 .06.2026 1803LarreyDJ . Автор описывает экзартикуляцию в тазобедренном суставе и методику пересечения LCF , которую именует «межсуставная связка». 16 .06.2026 1948EpsteinI . Реда ктор комментирует слова раввина Самуила (Шмуэля) в трактате Хуллин Вавилонского талмуда, поясняющего расположение LCF по отношению к суставу и ее отличие от седалищного нерва. 1753AstrucJ . Автор, анализируя книгу «Бытие» приводит текст на французском языке с упоминанием травмы тазобедренного сустава, повреждении LCF и последствиях инцидента. 29 .05.2026 Публикации о LCF в 2026 году (Май). Статьи и к ниги с упоминанием LCF опубликованные в мае 2026 года. 28 .05.2026 Интернет-журнал "О КРУГЛОЙ СВЯЗКЕ БЕДРА", май 2026 26 .05.2026 20c.Wikstrom B . Скульптура. Изображение обстоятельств и механ...

Primates: Prosimii & Anthropoidea
В позднем девоне или 367.5 млн. лет назад от земноводных произошла ветвь амниот (Amniota) (2011PyronRA). В той когорте изначально возникают синапсиды (Synapsid), потом зауропсиды (Sauropsida), а затем рептилии (Reptilia) (1995LaurinM_ReiszRR). Первые представители рептилообразных (Reptilomorpha) четвероногих осваивали континенты 363-290 млн. лет назад (1995LundbergJG).
Самым ранним синапсидом признан Asaphestera platyris из раннего пенсильванского подпериода, длившегося с отметки 323.2±0.4 по 315.2±0.2 млн. лет назад (2020MannA_AndersonJS; 2023CohenKM_CarN). По данным молекулярных часов, млекопитающие (Mammalia), именуемые также звери (Theria), отмежевываются 310 млн. лет назад от побега, протянувшегося к рептилиям (2004HedgesSB_ShoeJL). Первые вскармливающие детенышей молоком животные выделились среди примитивных синапсид – предков цинодонтов (Cynodontia) (2013VaughanTA_CzaplewskiNJ). Старейшего млекопитающего Liaoconodon hui нашли в отложениях начала мелового периода, то есть жившего 145.0-100.5 млн. лет назад (2011MengJ_LiC; 2023CohenKM_CarN).
Молекулярные
часы «показывают»: надотряд эуархонтоглиры (Euarchontoglires) обособились в классе
млекопитающих 88.8 млн. лет назад (2007JaneckaJE_MurphyWJ). Эта систематическая
единица, также называемая надприматы (Supraprimates), подразделяется на:
грандотряд грызунообразные (Glires) с отрядами зайцеобразные (Lagomorpha)
и грызуны (Rodentia); грандотряд эуархонты (Euarchonta) с отрядами тупайи (Scadentia)
и миротрядом приматообразные (Primatomorpha), в котором различают отряды:
шерстокрылы (Dermoptera) и приматы (Primates = «Вожди творения») (2017EsselstynJA_FairclothBC).
 |
| Светлолобая коата; экспонат Зоологического музея МГУ (Москва), фотография автора. |
По молекулярным
свидетельствам, эуархонты формируются 87.9 млн. лет назад, приматообразные – 86.2
млн. лет назад, приматы – 79.6 млн. лет назад, а тупайи – 63.4 млн. лет назад (2007JaneckaJE_MurphyWJ). В соответствии с летописью окаменелостей, плацентарные млекопитающие
диверсифицировались примерно 66 млн. лет назад, а самый ранний
плезиадапиформный примат жил 65 млн. лет назад (2015ChesterSG_ClemensWA). Древнейшие останки несомненного примата Teilhardina asiatica изысканы
в пластах почвы начала эоцена или возрастом 55.5 млн. лет (2006SmithT_GingerichPD; 2023CohenKM_CarN).
Обезьяны
обозначились в мире животных приблизительно 50 млн. лет назад (2009SrivastavaRP). Один из исходных гоминоидов
Morotopithecus bishop процветал в Африке 20.6
млн. лет назад (1997GeboDL_PilbeamD). Его близкий «родственник»
примат Afropithecus жил 17.5-17.0 млн. лет назад (1997LeakeyM_WalkerA). По молекулярным оценкам, орангутаны дифференцируются 19.3-15.7 млн.
лет назад, гориллы – 9.7-7.6 млн. лет назад, а шимпанзе – 6.5-5.8 млн. лет
назад (2011IsrafilH_SteiperME). Эволюционисты полагают:
обособление гиббонов состоялось 29.62-20.68 млн. лет назад, орангутанов – 18.42-12.53
млн. лет назад, горилл – 9.89-6.62 млн. лет назад, обыкновенных шимпанзе – 6.52-4.77
млн. лет назад, карликовых шимпанзе (бонобо) – 5.85-4.35 млн. лет назад (2022PoszewieckaB_GambinA).
Сейчас
в отряде приматов насчитывается 190 видов, помещаемых в подотряд низшие приматы
(Prosimii) с семействами: тупаииды (Tupaiidae), лори (Lorisidae),
лемуры (Lemuridae); и в подотряд высшие приматы (Anthropoidea) с семействами: цепкохвостые обезьяны или капуцины (Cebidae),
мартышки (Cercopithectidae), человекообразные обезьяны (Pongidae)
и гоминиды (Hominidae) с единственным видом человек разумный (Homo sapiens) (1979НаумовНП_КарташевНН). В Африке и Азии до сих
пор здравствуют семь видов человекообразных обезьян трех родов: орангутан (Pongo),
горилла (Gorilla) и шимпанзе (Pan) (2005WilsonDE_ReederDM). Ныне орангутаны и гориллы составляют подсемейство гоминины (Homininae),
а человек и шимпанзе объединены в трибу гоминини (Hominini), как произошедших от
общего предка (2001GrovesCP).
О наличии ligamentum capitis femoris (LCF) у животного можно судить на основании анализа вертлужной впадины и проксимального отдела бедра. На бедренной кости – это ямка головки бедренной кости, борозда головки бедренной кости, бугристость или щель на головке бедренной кости, краевой дефект суставной поверхности головки бедренной кости. В области таза о присутствии LCF свидетельствуют: вырезка вертлужной впадины, ямка вертлужной впадины, отверстие дна вертлужной впадины, неровности на суставной поверхности вертлужной впадины.
Как нам удалось выяснить, тема
наличия LCF у человекообразных обезьян
впервые заинтересовала исследователей в 19-м веке. Отмечались случаи отсутствия
данной структуры и велись жаркие дискуссии по этому вопросу. R. Owen (1835) обнаружил
углубление на головке бедренной кости для LCF у шимпанзе,
однако не нашел ее признаков у орангутана. G. Mivart (1869) выявил лишь в одном
скелете орангутана ямку головки бедренной кости, а также иногда не находил
следов LCF у
горилл. E. Moser (1893) отмечает, что у орангутана LCF обычно
отсутствует. Морфологическое исследование E.S. Crelin (1988) взрослого самца орангутана позволило
визуализировать LCF, прикрепленную к головке бедренной
кости.
В настоящее время установлено,
что все человекообразные обезьяны имеют LCF. Ярким признаком ее присутствия и функционирования
при жизни является отчетливая ямка вертлужной впадины.
 |
| Горилла. Левая вертлужная впадина (вид снаружи); 3-D модель таза взрослой самки гориллы (9.8 лет) зоопарка Барселоны. [sketchfab.com] |
 |
| Шимпанзе. Правая вертлужная впадина (вид снаружи), таз взрослой особи шимпанзе; экспонат Палеонтологического музея им. Ю.А. Орлова (Москва), фотография автора. |
 |
| Орангутан. Правая вертлужная впадина (вид снаружи); 3-D модель бедренной кости мужской особи взрослого борнейского орангутана зоопарка Барселоны. [sketchfab.com] |
В отдельных
случаях LCF оставляет след своего крепления непосредственно на
суставной поверхности головки бедренной кости.
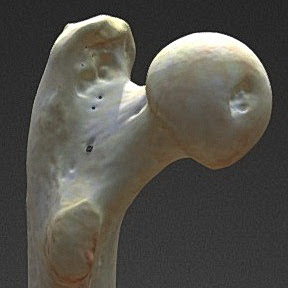 |
| Горилла. Проксимальный отдел левой бедренной кости (вид сзади); 3-D модель бедренной кости взрослой самки гориллы (9.8 лет) зоопарка Барселоны. [sketchfab.com] |
 |
| Шимпанзе. Проксимальный отдел правой бедренной кости (вид сзади); 3-D модель бедренной кости шимпанзе из учебной коллекции Исследовательских лабораторий археологии Университета Северной Каролины в Чапел-Хилл (модель Стива Дэвиса). [sketchfab.com] |
Иногда местом дистального прикрепления является край головки бедренной кости, причем у разных видов.
 |
| Орангутан. Проксимальный отдел правой бедренной кости (вид сзади); 3-D модель бедренной кости мужской особи взрослого борнейского орангутана зоопарка Барселоны. [sketchfab.com] |
 |
| Горилла. Левая тазовая кость и проксимальный отдел бедренной кости (отведение, супинация); экспонат Зоологического музея МГУ (Москва), фотография автора. |
Использованная литература
Cohen KM, Harper
DAT, Gibbard PL, Car N. The International Commission on Stratigraphy (ICS)
International Chronostratigraphic Chart. September 2023. [stratigraphy.org]
Pyron RA.
Divergence time estimation using fossils as terminal taxa and the origins of
Lissamphibia. Systematic biology. 2011;60(4)466-81. [academic.oup.com]
Laurin M, Reisz RR.
A reevaluation of early amniote phylogeny. Zoological Journal of the Linnean
Society. 1995;113(2)165-223. [academia.edu]
Lundberg JG.
Chordata. The Tree of Life Web Project. Version 1 January 1995 (under
construction). [tolweb.org]
Mann A, Gee BM,
Pardo JD, Marjanović D, Adams GR, Calthorpe AS, Maddin HC, Anderson JS.
Reassessment of historic ‘microsaurs’ from Joggins, Nova Scotia, reveals hidden
diversity in the earliest amniote ecosystem. Papers in Palaeontology.
2020;6(4)605-25. [researchgate.net]
Hedges SB, Blair
JE, Venturi ML, Shoe JL. A molecular timescale of eukaryote evolution and the
rise of complex multicellular life. BMC evolutionary biology. 2004;4(1)1-9. [link.springer.com]
Vaughan TA, Ryan
JM, Czaplewski NJ. Mammalogy. Sudbury: Jones & Bartlett Learning, 2013. [books.google]
Meng J, Wang Y, Li
C. Transitional mammalian middle ear from a new Cretaceous Jehol eutriconodont.
Nature. 2011;472(7342)181-5. [researchgate.net]
Janecka JE, Miller
W, Pringle TH, Wiens F, Zitzmann A, Helgen KM, Springer MS, Murphy WJ.
Molecular and genomic data identify the closest living relative of primates.
Science. 2007;318(5851)792-4. [epository.si.edu]
Esselstyn JA,
Oliveros CH, Swanson MT, Faircloth BC. Investigating difficult nodes in the
placental mammal tree with expanded taxon sampling and thousands of
ultraconserved elements. Genome Biology and Evolution. 2017;9(9)2308-21. [scholar.google]
Chester SG, Bloch
JI, Boyer DM, Clemens WA. Oldest known euarchontan tarsals and affinities of
Paleocene Purgatorius to Primates. Proceedings of the National Academy of
Sciences. 2015;112(5)1487-92. [scholar.google]
Smith T, Rose KD,
Gingerich PD. Rapid Asia-Europe-North America geographic dispersal of earliest
Eocene primate Teilhardina during the Paleocene-Eocene Thermal Maximum. Proc
Natl Acad Sci USA. 2006;103:11223-7. [scholar.google]
Srivastava RP.
Morphology of the Primates and Human Evolution. New Delhi: PHI Learning Pvt. Ltd.,
2009. [books.google]
Gebo DL, MacLatchy
L, Kityo R, Deino A, Kingston J, Pilbeam D. A hominoid genus from the early
Miocene of Uganda. Science. 1997;276:401-4. [researchgate.net]
Leakey M, Walker A.
Afropithecus: function and phylogeny. In: Begun DR, Ward CV, Rose MD (Eds).
Function, phylogeny and fossils: Miocene hominoid evolution and adaptations.
New York: Plenum, 1997:225-39. [link.springer.com]
Israfil H, Zehr SM,
Mootnick AR, Ruvolo M, Steiper ME. Unresolved molecular phylogenies of gibbons
and siamangs (Family: Hylobatidae) based on mitochondrial, Y-linked, and
X-linked loci indicate a rapid Miocene radiation or sudden vicariance event.
Molecular Phylogenetics and Evolution. 2011;58(3)447-55. [ncbi.nlm.nih.gov]
Poszewiecka B,
Gogolewski K, Stankiewicz P, Gambin A. Revised time estimation of the ancestral
human chromosome 2 fusion. BMC genomics. 2022;23(6)1-16. [link.springer.com]
Наумов НП, Карташев НН. Зоология позвоночных. Ч. 2.
Пресмыкающиеся, птицы, млекопитающие: Учебник для биолог. спец. ун-тов. Москва:
Высшая школа, 1979. [chembaby.ru]
Wilson DE, Reeder
DM (Eds). Mammal species of the world: a taxonomic and geographic reference.
Vol. 1. Baltimore: Johns Hopkins University Press, 2005. [books.google]
Groves CP. Towards
a taxonomy of the Hominidae. In: Humanity from African Naissance to Coming Millennia.
In: Tobias PV, Raath MA, Moggi-Cecchi J, Doyle GA (Eds). Colloquia in Human
biology and Palaeonthropology. Firenze: Firenze University Press, 2001:291-7. [library.oapen.org]
Crelin ES. Ligament of the head of the femur in the orangutan and indian elephant. The Yale J Biol Med. 1988;61(5)383-8. [ncbi.nlm.nih.gov, ncbi.nlm.nih.gov.pdf]
Owen R. On the osteology of the Chimpanzee and Orang. Transactions of the Zoological Society of London. Vol. I. London, 1835:343-379. [books.google]
Mivart G. Contributions towards a more complete knowledge of the Skeleton of the Primates. Part I. The Appendicular Skeleton of Simia. Transact. Zool. Soc. 1869;6:175-226. [biodiversitylibrary.org]
Moser E. Ueber das Ligamentum teres des Hüftgelenks. Morphologische Arbeiten. 1893;2(1)36-92. [books.google , jstor.org]
Ключевые слова:
ligamentum capitis femoris, ligament of head of femur, ligamentum teres, круглая связка, связка головки бедра, палеонтология, эволюция, животные, обезьяна
NB! Добросовестная практика использования: копирование для целей критики, обзора, комментариев, исследований и частного изучения в соответствии с Законами об авторском праве: Copyright Laws of the US: 17 U.S.C. §107; Copyright Law of the EU: Dir. 2001/29/EC, art.5/3a,d; Copyright Law of the RU: ГК РФ ст.1274/1.1-2,7.